


2.T细胞受体
T细胞受体(TCR)是T细胞表面的分子,作为细胞对外界的“眼睛”。没有这些受体,T细胞就会盲目飞行,无法感知外界的情况。T细胞受体有两种类型:αβ和γδ。每种类型的受体由两种蛋白质组成,分别是α和β或γ和δ。与B细胞受体的重链和轻链一样,α、β、γ和δ的基因是通过混合和匹配基因片段来组装的。事实上,在B细胞和T细胞中,相同的蛋白质(RAG1和RAG2)通过在染色体DNA中制造双链断裂来启动基因片段的拼接。当基因片段混合和匹配时,会展开一场“竞争”,每个T细胞最终会带有αβ或γδ受体,但不会同时带有两者。通常,成熟T细胞上的所有TCR都是相同的——尽管这一规则也有例外。
2.1.传统T细胞
2.2.非传统T细胞
3.T细胞受体如何传递信号
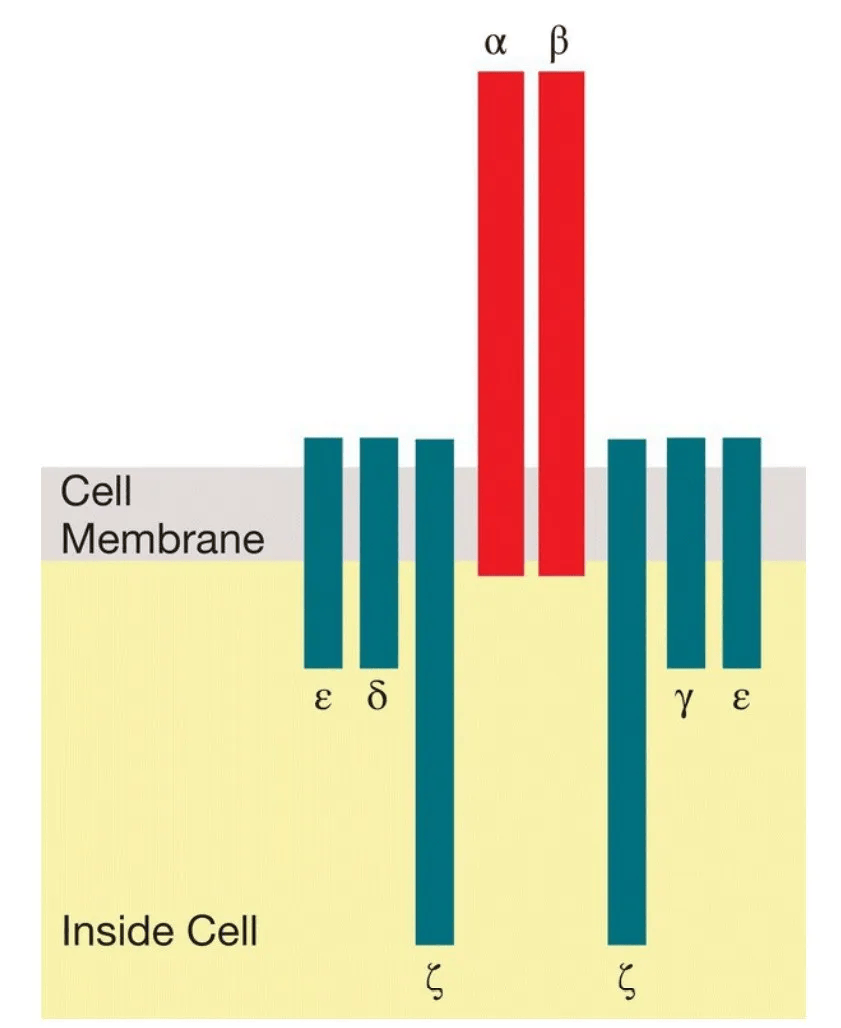
4.CD4和CD8共受体
当T细胞开始在胸腺成熟时,它们在表面表达两种类型的共受体。免疫学家称它们为CD4+CD8+或“双阳性”细胞。重要的是,CD4共受体只会“夹在”类II MHC分子上,CD8共受体只会与类I MHC分子匹配。因此,CD4和CD8共受体有助于集中Th细胞和CTL对适当的MHC分子的注意力。最新的观点是,双阳性T细胞扫描APC,寻找它们的TCR可以结合的MHC分子。如果T细胞的受体形状正确,可以结合APC表面上的类II MHC分子,CD4共受体就会夹上。相反,如果TCR形状正确,可以结合类I MHC分子,CD8共受体就会连接。

CD4和CD8分子的尾部穿过T细胞的质膜并延伸到细胞质中。尽管这些尾部不同,但它们都有发出信号的能力。因此,当CD4分子夹在类II MHC分子上时,CD8共受体的表达会被下调,T细胞变成了一个“单阳性”的CD4+ T细胞,承诺作为辅助T细胞发挥作用。相比之下,如果CD8共受体夹在类I MHC分子上,CD4的表达会被终止,那个细胞就变成了一个杀手T细胞。这就是理论,但这些共受体分子如何帮助“指导”T细胞作为辅助细胞或杀手细胞发挥作用的具体机制尚不清楚。
5.共刺激
在初始T细胞中,细胞受体与其细胞核之间的“连接”不是很好。就好像T细胞有一个电气系统,在传感器(TCR)和它设计用来调节的设备(细胞核中的基因表达)之间放置了一个大电阻器。由于这个“电阻器”,TCR发出的信号在传递到细胞核的过程中会丢失很多。结果是,需要大量TCRs与它们相应的抗原结合,才能使到达细胞核的信号足够强烈以产生任何效果。然而,如果TCRs在结合的同时,T细胞也接收到共刺激,TCRs发出的信号会被放大很多倍,因此需要较少的(可能大约少100倍)TCRs结合才能激活一个初始T细胞。
尽管已经鉴定出许多可以共刺激T细胞的不同分子,但研究得最好的例子是B7蛋白(B7-1和B7-2),它们表达在抗原呈递细胞的表面。B7分子通过插入T细胞表面称为CD28的受体分子,为T细胞提供共刺激。
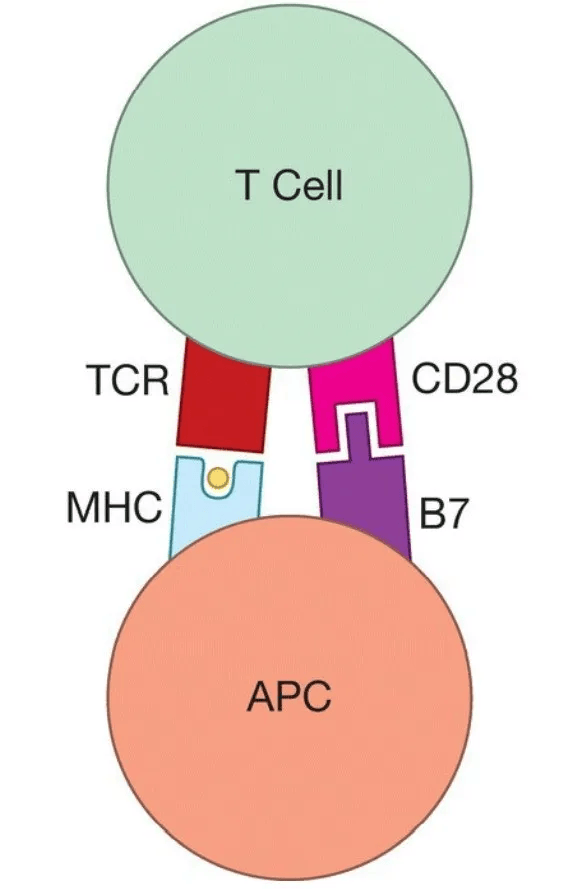
除了T细胞受体被MHC-肽复合物结合外,初始T细胞还必须接收共刺激信号才能被激活。共刺激可以被看作是一个“放大器”,它加强了T细胞受体发出的“我已参与”信号,从而降低了必须由MHC-肽复合物交联的TCRs的阈值数量。有趣的是,一旦初始T细胞被激活,TCRs与细胞核之间的联系就会加强。就好像一个有经验的T细胞已经被“重新布线”,使得初始T细胞中存在的电阻被绕过了。由于这种重新布线,TCR信号的放大对于有经验的T细胞来说不像对原始T细胞那么重要。因此,有经验的T细胞对共刺激的需求减少了。
6.时间延时摄影:辅助T细胞的激活过程
在淋巴结中,辅助T细胞快速扫描树突状细胞,以查看它们的同源抗原是否被展示。一个树突状细胞通常每小时接待约1000次这样的“访问”。如果T细胞发现一个展示其同源抗原的树突状细胞,T细胞会“徘徊”,因为初始辅助T细胞的完全激活通常需要几个小时。在此期间,许多重要事件发生。首先,树突状细胞表面的粘附分子与其在T细胞上的粘附伙伴结合,帮助将两个细胞保持在一起。接下来,T细胞表面的CD4共受体分子夹在树突状细胞的II类MHC分子上,加强了两个细胞之间的相互作用。此外,其TCRs的参与上调了Th细胞表面的粘附分子表达,加强了将APC和T细胞固定在一起的“胶水”。这很重要,因为TCR与MHC-肽复合物之间的初始结合实际上相当弱——以允许快速扫描。因此,像Velcro一样的粘附分子对T细胞激活极为重要。TCRs和粘附分子在APC和T细胞接触点的聚集导致了免疫学家所说的免疫突触的形成。
辅助T细胞受体的参与还上调了其表面CD40L蛋白的表达,当这些蛋白插入树突状细胞表面的CD40蛋白时,会发生几件了不起的事情。尽管成熟的树突状细胞在最初进入淋巴结时就表达MHC和共刺激分子(例如,B7),但当APC上的CD40蛋白被Th细胞上的CD40L蛋白参与时,这些蛋白的表达水平会增加。此外,树突状细胞的CD40蛋白的参与延长了树突状细胞的寿命。这种“有用”树突状细胞寿命的延长是有意义的。它确保了展示T细胞同源抗原的特定树突状细胞会停留足够长的时间,以帮助激活许多这些T细胞。因此,树突状细胞与初始辅助T细胞之间的相互作用不仅仅是单向的。这些细胞实际上进行了一种激活“舞蹈”,它们相互刺激。这种合作的最终结果是,树突状细胞变成了一个更有效的抗原呈递细胞,而Th细胞被激活以表达帮助激活B细胞所需的高水平CD40L。
因此,辅助T细胞激活过程中的事件顺序如下:粘附分子介导Th和APC之间的弱结合,而TCRs参与其由APC呈递的同源抗原。TCR参与加强了两个细胞之间的粘附,并上调了Th细胞上CD40L的表达。然后CD40L与APC上的CD40结合,并增加了APC表面上MHC和共刺激分子(例如,B7)的表达。APC向Th细胞提供的共刺激放大了“TCR参与”信号,导致Th细胞的激活。因此,由于B7/CD28和CD40L/CD40相互作用的结果,Th细胞和DC实际上相互刺激。对他们双方来说都是双赢的。

7.杀手T细胞是如何被激活的?
对于原始Th细胞和CTL的激活只需要两个细胞的相互作用,在启动适应性免疫系统以在入侵者完全接管之前是完全合理的。然而,尽管没有Th细胞帮助激活的CTL也会增殖一些以增加它们的数量并且可以杀死感染的细胞,但这些“无助”的CTL不会高效地杀死,并且它们的寿命不长。因此,CTL的无助激活导致一小群杀手T细胞旨在快速应对感染初期的入侵者。相比之下,得到辅助性T细胞帮助完全激活的CTL可以健壮地增殖,可以高效地杀死,并且可以成为记忆杀手T细胞(可以防御同一攻击者的后续入侵的细胞)。当然,这又回到了CTL如何能够在不需要三个细胞相互作用的情况下通过Th细胞和DC完全激活的问题。
一种可能性是“顺序模型”,该模型假设当辅助性T细胞被激活时,激活它们的树突状细胞变得“有资格”完全激活CTL。根据这一模型,DC的CD40蛋白与Th细胞的CD40L蛋白的结合改变了(授权)DC,使其在与Th细胞脱离后,能够完全激活随后接触到的CTL。人们认为这种授权涉及DC表面蛋白的上调,然后可以与CTL表面的受体蛋白结合,但这并不确定。在这个顺序模型中,DC和Th细胞首先相遇,然后授权的DC和CTL稍后相遇,这将避免所有三个细胞必须同时相遇的需要。在这个场景中,Th细胞通过使用树突状细胞作为中介间接帮助激活CTL。一旦CTL被完全激活并前往战场,Th细胞可以直接帮助CTL,通过提供如IL-2这样的细胞因子以保持CTL的全力运作。
CTL是如何被激活的是一个优秀的例子,这个问题尚未被免疫学家完全回答。而这是一个重要的问题!了解CTL是如何被激活的对于设计疫苗和制定涉及CTL的疾病的治疗方案至关重要。
回顾
对于B细胞或T细胞的激活,它们的受体必须被抗原聚集,因为这种交联将许多信号分子汇集在细胞的一个小区域内。当信号分子的密度足够大时,就会引发一个酶促链反应,将“受体参与”信号传递到细胞核。在那里,细胞的“大脑中心”,激活相关的基因会因这个信号而被关闭或开启。
尽管受体的交联对于B细胞或T细胞的激活至关重要,但这还不够。原始的B细胞和T细胞还需要非抗原特异性的共刺激信号。这种双重信号需求为激活提供了一个安全系统,防止B细胞或T细胞的不当激活。对于B细胞的激活,辅助性T细胞可以通过表面蛋白CD40L提供共刺激,这些蛋白与B细胞表面的CD40蛋白结合。B细胞也可以通过“危险信号”共刺激,包括入侵者特异性的分子标志或战斗细胞因子。
对于T细胞来说,共刺激通常涉及激活的树突状细胞上的B7蛋白,这些蛋白与T细胞表面的CD28蛋白结合。
在感染初期,B细胞和杀手T细胞可以在没有辅助性T细胞的帮助下被激活。无助的浆B细胞产生IgM抗体,因为它们没有转换到可能更适合防御特定入侵者的抗体类别。它们通常没有经历过体细胞超突变,所以它们的BCRs没有被“微调”。它们只活很短的时间。同样,无助的CTLs不会大量增殖,寿命短,并且没有得到辅助性T细胞帮助的T细胞那样高效地杀死。尽管无助的B细胞和T细胞有这些缺陷,但它们可以在更“复杂”的B细胞和T细胞被产生的同时,为病原体提供重要的快速反应。
B细胞和T细胞之间的另一个区别是,在感染期间,BCR可以经历体细胞超突变和选择。所以B细胞可以从牌堆中“抽牌”以尝试获得更好的牌。相比之下,TCR不会超突变,所以T细胞必须满足于它们手中的牌。返回搜狐,查看更多


